Sat, Jul 12, 2025
Volume 19, Issue 1 (Winter & Spring 2022)
ASJ 2022, 19(1): 39-46 |
Back to browse issues page

Download citation:
BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:



BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:
Ehi Oche P, Isaac Dibal N, Chiroma S M, Orendu Oche Attah M. Morphological and Morphometric Variations of Erythrocytes in Clarias Gariepinus, Bufo Bufo, and Agama Agama Using Some Histological Stains. ASJ 2022; 19 (1) :39-46
URL: http://anatomyjournal.ir/article-1-300-en.html
URL: http://anatomyjournal.ir/article-1-300-en.html



1- Department of Human Anatomy, Faculty of Basic Medical Sciences, University of Maiduguri, Nigeria.
Full-Text [PDF 3498 kb]
(648 Downloads)
| Abstract (HTML) (1845 Views)
Full-Text: (1006 Views)
1. Introductio
Erythrocytes are highly specialized and the most abundant cell type in vertebrates’ blood. Their primary function is the transportation of oxygen to tissues of the body via hemoglobin [1, 2]. Except for ice fishes, all vertebrates have hemoglobin packed in their erythrocytes. Erythrocytes are used in the eco-physiological and eco-morphological evaluation of species [3, 4]. Blood cell parameters are used to evaluate the physiological and health status of individuals of the same specie [5]. Blood parameters such as volume, fragility, hematocrit, pH, number, and sizes may vary in different species and even among the same species for some reasons [6]. Factors that affect blood parameters in the same species include differences in altitude, body mass, age, sex, and diseases such as sickle cell and megaloblastic anemia [7, 8]. The variation in the blood parameters of different species results from divergent evolutionary changes [9].
Stains are used in histology to highlight tissue characteristics and to enhance contrast; they are employed based on their affinity to different tissues [10, 11]. Staining is a procedure undertaken in histological technique for studying tissue with a microscope; it involves introducing dyes into tissues for better visualization [12]. Staining has various applications in biological and medical sciences, including demonstrating certain substances within tissues and diagnosing different diseases [13]. The most commonly used histological stains include hematoxylin, eosin, methylene blue, cresyl violet, Giemsa, periodic acid Schiff (PAS), Leishman, and Papanicalau [14, 15, 16].
This study aimed to evaluate the affinity of different histological stains; Giemsa, Hematoxylin and eosin (H&E), and methylene blue on the erythrocytes of Clarias gariepinus (Catfish), Bufo bufo (Toad) and Agama agama (Lizard). Additionally, the erythrocytes morphology of stains with the best affinity were described and schematically drawn besides their morphometric analysis.
2. Materials and Methods
Experimental animal
Three of the following vertebrates were used for the study (C. gariepinus, B. bufo, and A. agama). C. gariepinus and B. bufo were obtained from Lake Alau in Maiduguri, Nigeria, while the A. agama was obtained from the University of Maiduguri campus. The research was approved by the Department of Human Anatomy ethical committee (Code: UM/HA/UGP19.20-095) and conducted following the National Research Council Guide for the use of laboratory animals (8th edition).
Smear preparation and staining
The blood sample of each animal was collected using a syringe. At least 10 smears from each animal were made on glass slides at different intervals and stained with hematoxylin (C16H14O6) and eosin (C20H6Br4Na2O5), Giemsa stain (mixture of buffered Azure B, methylene blue and eosin Y in methanol and glycerol) and methylene blue (C16H18ClN3S). The stains were left for 5-10 minutes each, washed off under running water, and then the slides were observed under a light microscope (Leica Biosystems RM2235, Germany). Photomicrographs were taken at x400 magnification (40 objectives x10 oculars) using a microscope camera (AmScope, M500, USA).
Morphometric study
The diameters of the erythrocytes were measured using a standard ocular micrometer from the stains that produced the best contrast. The erythrocyte’s diameter of an oval cell is the length of the short axis (Figure 1A), while the diameter of a spherical cell is the length of a straight line passing through the center of the cell, whose endpoints lie on the cell membrane (Figure 1B).
.jpg)
The area and volume of each cell were calculated using the following Equation:
Area of spherical, area of oval cell=πab, the volume of spherical cell=(4/3π)r3, the volume of oval cell=(4/3π)a2b
, where r is the radius, r=(diameter/2), a is the radius of the long axis, a=(long axis/2), b is the radius of the short axis, b=(short axis/2) (Figure 1C), π=3.142.
Statistical analysis
Data were analyzed with GraphPad Prism 7 (GraphPad Software, California, USA). A schematic diagram of best-stained cells was drawn. One-way analysis of variance followed by the Bonferroni post-hoc test was used to compare the erythrocytes’ diameter of different vertebrates, and values were expressed as Mean±SEM. P<0.05 was considered statistically significant.
3. Results
Morphology
The erythrocytes of C. gariepinus are spherical, as seen on Giemsa, H&E, and methylene blue stained slides. The nuclei are stained violet with Giemsa, pink with H&E, and blue with methylene blue (Figures 2, 3, 4). The cell membranes were distinguishable from the background in all stains but distinct in Giemsa stained cells compared to H&E and methylene blue stained cells (Figures 2, 3, 4). The cytoplasm showed less affinity for all the stains compared to their nuclei. The cytoplasm of erythrocytes in C. gariepinus had a higher affinity for Giemsa stain when compared to H&E and methylene blue (Figures 2, 3, 4)..jpg)
.jpg)
.jpg) Erythrocytes of B. bufo range from spherical to oval shapes, as demonstrated by Giemsa, H&E, and methylene blue stains (Figures 5, 6, 7). With Giemsa, the nuclei appeared dark blue with orchid cytoplasm, H&E stain highlighted blue nuclei with pink cytoplasm, while methylene blue stained cells showed a cytoplasm that was lightly stained blue with dark nuclei (Figures 5, 6, 7). Giemsa demonstrated B. bufo erythrocytes better than H&E and methylene blue stains.
Erythrocytes of B. bufo range from spherical to oval shapes, as demonstrated by Giemsa, H&E, and methylene blue stains (Figures 5, 6, 7). With Giemsa, the nuclei appeared dark blue with orchid cytoplasm, H&E stain highlighted blue nuclei with pink cytoplasm, while methylene blue stained cells showed a cytoplasm that was lightly stained blue with dark nuclei (Figures 5, 6, 7). Giemsa demonstrated B. bufo erythrocytes better than H&E and methylene blue stains.


 The erythrocytes of A. agama are oval-shaped with purple nuclei and pink cytoplasm when stained with H&E (Figure 8).
The erythrocytes of A. agama are oval-shaped with purple nuclei and pink cytoplasm when stained with H&E (Figure 8).
 Giemsa and methylene blue stained cells showed no contrast between the nucleus and cytoplasm. The whole cell appeared blue in both Giemsa and methylene blue-stained cells (Figures 9, 10). H&E stain demonstrated the erythrocytes of A. agama better when compared to Giemsa and methylene blue stains.
Giemsa and methylene blue stained cells showed no contrast between the nucleus and cytoplasm. The whole cell appeared blue in both Giemsa and methylene blue-stained cells (Figures 9, 10). H&E stain demonstrated the erythrocytes of A. agama better when compared to Giemsa and methylene blue stains.

 Morphometry
Morphometry
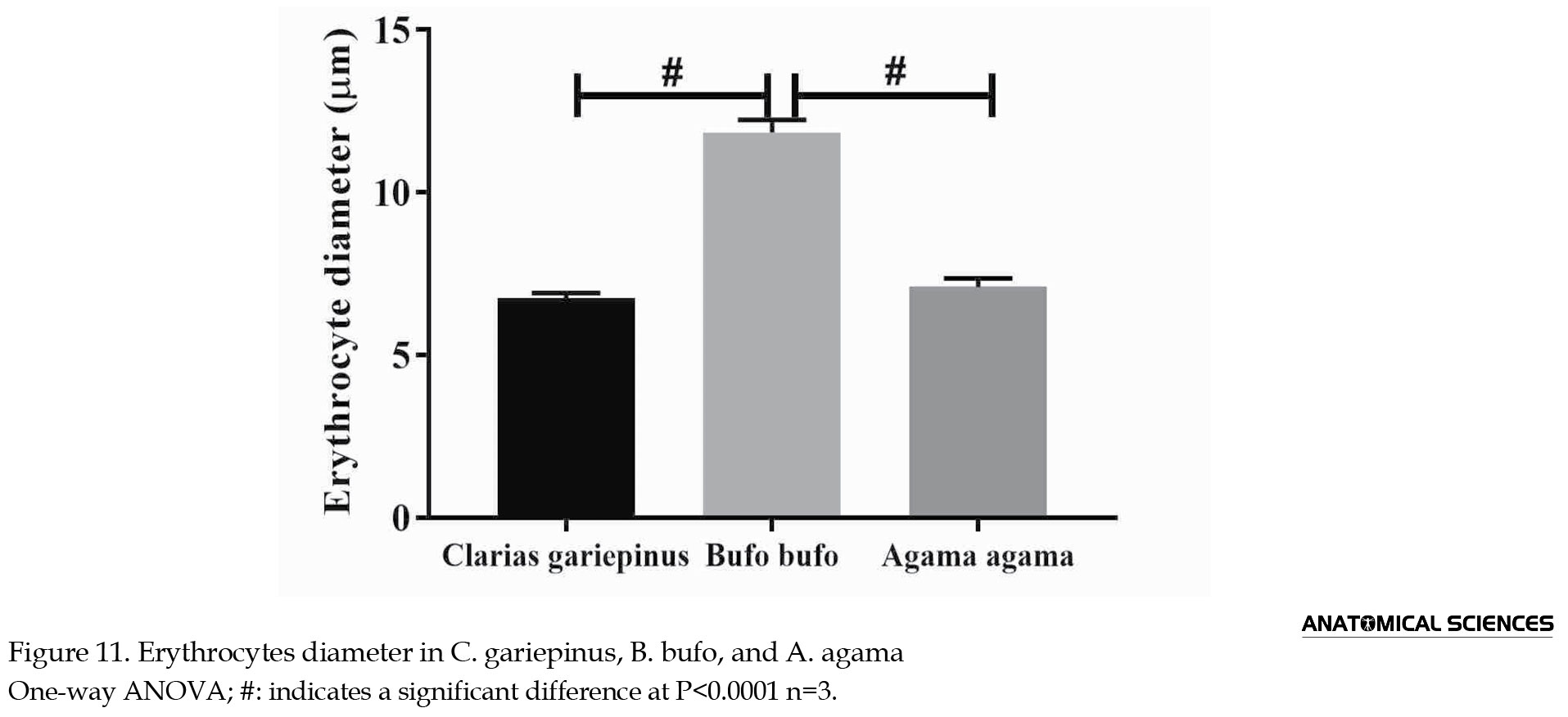
The erythrocyte’s diameter in C. gariepinus, B. bufo, and A. agama was 6.75±0.16 µm, 11.85±0.38 µm, and 7.10±0.26 µm, respectively. The diameter of B. bufo erythrocytes was significantly wider (P<0.001) compared to C. gariepinus; likewise, the B. bufo erythrocytes’ diameter was significantly wider (P<0.001) when compared to A. agama. However, there were no statistically significant differences between the erythrocytes diameter (P>0.05) in C. gariepinus and A. agama (Figure 11).
C. gariepinus had the least erythrocytes’ volume (36.17±1.73 µm3) compared to A. agama (61.50±2.66 µm3) and B. bufo (112.40±7.22 µm3). The erythrocytes’ area in B. bufo was significantly higher when compared to C. gariepinus and A. agama (P<0.001). C. gariepinus erythrocytes were significantly lower (P<0.001) compared to A. agama (Figure 12).

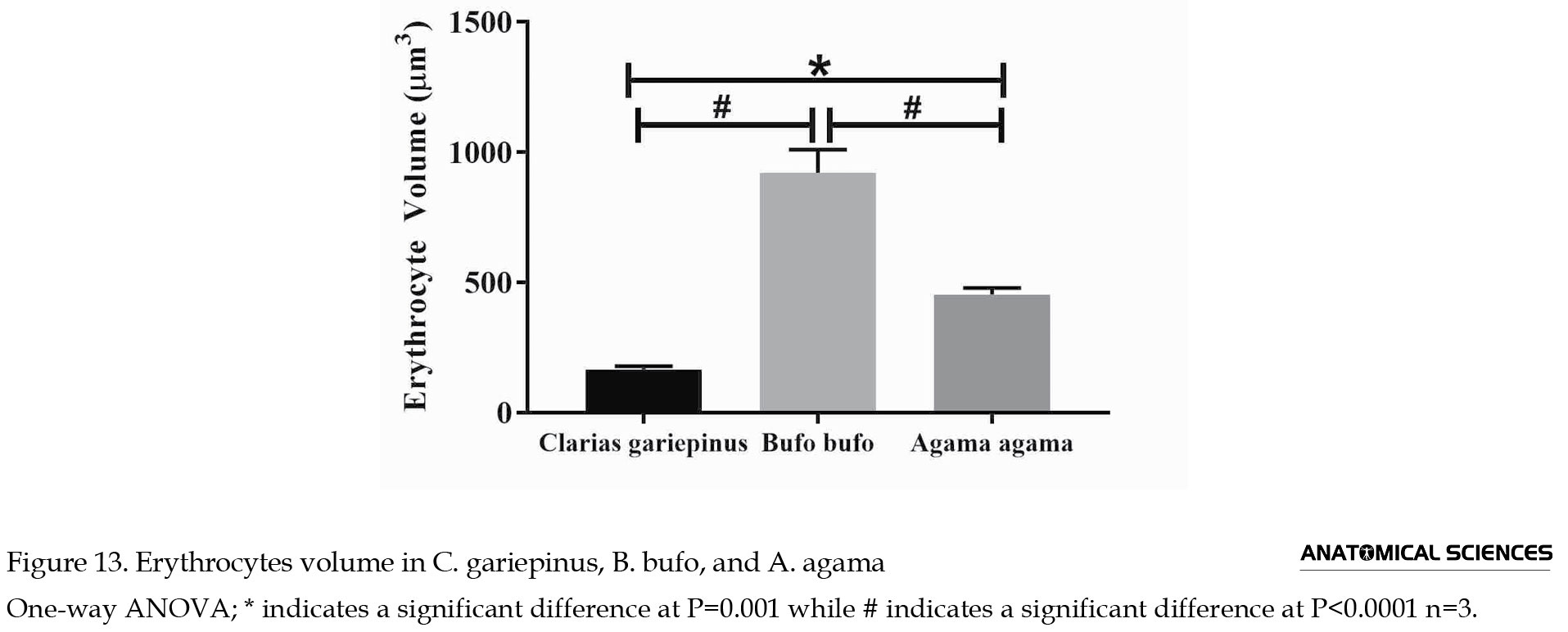
The volume of erythrocyte was 165.90±12.04 µm3 in C. gariepinus, 920.60±88.49 µm3 in B. bufo, and 453.60±24.51 µm3 in A. agama. The volume of the erythrocyte of A. agama was significantly higher (P=0.001) when compared to C. gariepinus. The volume of erythrocyte of B. bufo was significantly higher (P<0.001) compared to A. agama. The volume of the erythrocyte of C. gariepinus erythrocyte volume was significantly lower (P<0.001) when compared to B. bufo (Figure 13).
4. Discussion
In the current study, the erythrocyte’s shape ranges from spherical in C. gariepinus, oval to spherical in B. bufo, and oval in A. agama. Erythrocytes’ shapes vary greatly in fishes, from oval in tilapia and Channa punctata [17, 18] to elliptical in (Acipenser sinensis) Chinese sturgeon [19]. In triploid species of C. gariepinus, erythrocytes are elliptical, while diploid C. gariepinus are round in shape [20]. In general, the erythrocytes of fishes vary from oval to elliptic, with few round shapes [21]. In amphibians, the erythrocyte’s shape also ranges from oval in Bufo gargarizans, Microphyla ornate, Rana esculenta, and Fejervarya limnocharis [6, 22] to round in Xenopus laevis [23] and elliptical in Bufo Vulgaris [24].
Urodeles were reported to have more elongated erythrocytes than Anurans [25]. Circular and oval-shaped erythrocytes are found in the tadpole of Microphyla ornate and Indian tree frog [26, 27]. Erythrocytes of A. agama were similar in shape to other lizard species, Gollotia simonyi machadoi and Leiolepsis belliana rubritaeniata [28, 29]. The lizards of the genus Podarcis, Alggyroides, Zootoca, Timon, and other reptiles, including the red-eared slider turtle, all possess oval-shaped erythrocytes [30, 31].
The current study showed that the erythrocytes of C. gariepinus, B. bufo, and A. agama have distinct nuclei. Mature erythrocytes of non-mammalian species retain nuclei and some organelles responsible for energy supply and protein synthesis [32, 33]. The organelles in the erythrocytes of non-mammalian vertebrates include mitochondria, lysosomes, ribosomes, and endoplasmic reticulum [18, 34].
The erythrocytes’ parameters (diameter, area, and volume) in B. bufo were significantly higher when compared with C. gariepinus and A. agama. The erythrocytes’ size was reported to have a direct association with chromosome number in fishes [19]. Increased erythrocytes’ size was noted with increasing chromosome number from diploid to triploid in C. gariepinus [20]. In vertebrates, erythrocytes’ size is significantly correlated with genome size [35, 36]. Among vertebrates, amphibians tend to have the largest erythrocytes size [37]. This was also observed in the current study. Amphibians were reported to have the largest genome size and the most significant variation in genome magnitude ranging from 0.95 pg in Platyplectrum ornatum to 140 pg in Necturus lewisi [38].
The large genome size in amphibians might cause a higher diameter, area and volume of erythrocytes in B. bufo compared with C. gariepinus and A. agama. Reptiles generally have a small genome size ranging from 1.1 pg to 5.4 pg [35]. Hence, the smaller erythrocytes size in A. agama compared with B. bufo. In fishes, erythrocyte sizes might differ in different species due to genome size. The erythrocytes of Australian lungfish with a genome magnitude of 50 pg were about 10-fold higher when compared with Siamese fighting fish with a genome size of 0.6 pg [35]. Therefore, genome size is essential in determining erythrocytes’ size in different vertebrates and even the same vertebrates in a different genus.
5. Conclusion
The current study indicated that C. gariepinus and B. bufo erythrocytes were better demonstrated with Giemsa stain while A. agama erythrocytes were better shown with H&E stain. This signifies that Erythrocytes’ affinity to stain varies in different vertebrates, probably due to environmental and genetic factors. The erythrocytes’ shape, size, area, and volume varies in different vertebrates, and the variation is associated with chromosome number and genome size.
Ethical Considerations
Compliance with ethical guidelines
The research was approved by the Department of Human Anatomy Ethical Committee (Code: UM/HA/UGP19.20-095) and conducted in accordance with National research council guide for the use of laboratory animals (8th edition).
Funding
This research did not receive any grant from funding agencies in the public, commercial, or non-profit sectors.
Authors' contributions
All authors equally contributed to preparing this article.
Conflict of interest
The authors declared no conflict of interest to declare.
References
Erythrocytes are highly specialized and the most abundant cell type in vertebrates’ blood. Their primary function is the transportation of oxygen to tissues of the body via hemoglobin [1, 2]. Except for ice fishes, all vertebrates have hemoglobin packed in their erythrocytes. Erythrocytes are used in the eco-physiological and eco-morphological evaluation of species [3, 4]. Blood cell parameters are used to evaluate the physiological and health status of individuals of the same specie [5]. Blood parameters such as volume, fragility, hematocrit, pH, number, and sizes may vary in different species and even among the same species for some reasons [6]. Factors that affect blood parameters in the same species include differences in altitude, body mass, age, sex, and diseases such as sickle cell and megaloblastic anemia [7, 8]. The variation in the blood parameters of different species results from divergent evolutionary changes [9].
Stains are used in histology to highlight tissue characteristics and to enhance contrast; they are employed based on their affinity to different tissues [10, 11]. Staining is a procedure undertaken in histological technique for studying tissue with a microscope; it involves introducing dyes into tissues for better visualization [12]. Staining has various applications in biological and medical sciences, including demonstrating certain substances within tissues and diagnosing different diseases [13]. The most commonly used histological stains include hematoxylin, eosin, methylene blue, cresyl violet, Giemsa, periodic acid Schiff (PAS), Leishman, and Papanicalau [14, 15, 16].
This study aimed to evaluate the affinity of different histological stains; Giemsa, Hematoxylin and eosin (H&E), and methylene blue on the erythrocytes of Clarias gariepinus (Catfish), Bufo bufo (Toad) and Agama agama (Lizard). Additionally, the erythrocytes morphology of stains with the best affinity were described and schematically drawn besides their morphometric analysis.
2. Materials and Methods
Experimental animal
Three of the following vertebrates were used for the study (C. gariepinus, B. bufo, and A. agama). C. gariepinus and B. bufo were obtained from Lake Alau in Maiduguri, Nigeria, while the A. agama was obtained from the University of Maiduguri campus. The research was approved by the Department of Human Anatomy ethical committee (Code: UM/HA/UGP19.20-095) and conducted following the National Research Council Guide for the use of laboratory animals (8th edition).
Smear preparation and staining
The blood sample of each animal was collected using a syringe. At least 10 smears from each animal were made on glass slides at different intervals and stained with hematoxylin (C16H14O6) and eosin (C20H6Br4Na2O5), Giemsa stain (mixture of buffered Azure B, methylene blue and eosin Y in methanol and glycerol) and methylene blue (C16H18ClN3S). The stains were left for 5-10 minutes each, washed off under running water, and then the slides were observed under a light microscope (Leica Biosystems RM2235, Germany). Photomicrographs were taken at x400 magnification (40 objectives x10 oculars) using a microscope camera (AmScope, M500, USA).
Morphometric study
The diameters of the erythrocytes were measured using a standard ocular micrometer from the stains that produced the best contrast. The erythrocyte’s diameter of an oval cell is the length of the short axis (Figure 1A), while the diameter of a spherical cell is the length of a straight line passing through the center of the cell, whose endpoints lie on the cell membrane (Figure 1B).
The area and volume of each cell were calculated using the following Equation:
Area of spherical, area of oval cell=πab, the volume of spherical cell=(4/3π)r3, the volume of oval cell=(4/3π)a2b
, where r is the radius, r=(diameter/2), a is the radius of the long axis, a=(long axis/2), b is the radius of the short axis, b=(short axis/2) (Figure 1C), π=3.142.
Statistical analysis
Data were analyzed with GraphPad Prism 7 (GraphPad Software, California, USA). A schematic diagram of best-stained cells was drawn. One-way analysis of variance followed by the Bonferroni post-hoc test was used to compare the erythrocytes’ diameter of different vertebrates, and values were expressed as Mean±SEM. P<0.05 was considered statistically significant.
3. Results
Morphology
The erythrocytes of C. gariepinus are spherical, as seen on Giemsa, H&E, and methylene blue stained slides. The nuclei are stained violet with Giemsa, pink with H&E, and blue with methylene blue (Figures 2, 3, 4). The cell membranes were distinguishable from the background in all stains but distinct in Giemsa stained cells compared to H&E and methylene blue stained cells (Figures 2, 3, 4). The cytoplasm showed less affinity for all the stains compared to their nuclei. The cytoplasm of erythrocytes in C. gariepinus had a higher affinity for Giemsa stain when compared to H&E and methylene blue (Figures 2, 3, 4).
The erythrocyte’s diameter in C. gariepinus, B. bufo, and A. agama was 6.75±0.16 µm, 11.85±0.38 µm, and 7.10±0.26 µm, respectively. The diameter of B. bufo erythrocytes was significantly wider (P<0.001) compared to C. gariepinus; likewise, the B. bufo erythrocytes’ diameter was significantly wider (P<0.001) when compared to A. agama. However, there were no statistically significant differences between the erythrocytes diameter (P>0.05) in C. gariepinus and A. agama (Figure 11).
C. gariepinus had the least erythrocytes’ volume (36.17±1.73 µm3) compared to A. agama (61.50±2.66 µm3) and B. bufo (112.40±7.22 µm3). The erythrocytes’ area in B. bufo was significantly higher when compared to C. gariepinus and A. agama (P<0.001). C. gariepinus erythrocytes were significantly lower (P<0.001) compared to A. agama (Figure 12).
The volume of erythrocyte was 165.90±12.04 µm3 in C. gariepinus, 920.60±88.49 µm3 in B. bufo, and 453.60±24.51 µm3 in A. agama. The volume of the erythrocyte of A. agama was significantly higher (P=0.001) when compared to C. gariepinus. The volume of erythrocyte of B. bufo was significantly higher (P<0.001) compared to A. agama. The volume of the erythrocyte of C. gariepinus erythrocyte volume was significantly lower (P<0.001) when compared to B. bufo (Figure 13).
4. Discussion
In the current study, the erythrocyte’s shape ranges from spherical in C. gariepinus, oval to spherical in B. bufo, and oval in A. agama. Erythrocytes’ shapes vary greatly in fishes, from oval in tilapia and Channa punctata [17, 18] to elliptical in (Acipenser sinensis) Chinese sturgeon [19]. In triploid species of C. gariepinus, erythrocytes are elliptical, while diploid C. gariepinus are round in shape [20]. In general, the erythrocytes of fishes vary from oval to elliptic, with few round shapes [21]. In amphibians, the erythrocyte’s shape also ranges from oval in Bufo gargarizans, Microphyla ornate, Rana esculenta, and Fejervarya limnocharis [6, 22] to round in Xenopus laevis [23] and elliptical in Bufo Vulgaris [24].
Urodeles were reported to have more elongated erythrocytes than Anurans [25]. Circular and oval-shaped erythrocytes are found in the tadpole of Microphyla ornate and Indian tree frog [26, 27]. Erythrocytes of A. agama were similar in shape to other lizard species, Gollotia simonyi machadoi and Leiolepsis belliana rubritaeniata [28, 29]. The lizards of the genus Podarcis, Alggyroides, Zootoca, Timon, and other reptiles, including the red-eared slider turtle, all possess oval-shaped erythrocytes [30, 31].
The current study showed that the erythrocytes of C. gariepinus, B. bufo, and A. agama have distinct nuclei. Mature erythrocytes of non-mammalian species retain nuclei and some organelles responsible for energy supply and protein synthesis [32, 33]. The organelles in the erythrocytes of non-mammalian vertebrates include mitochondria, lysosomes, ribosomes, and endoplasmic reticulum [18, 34].
The erythrocytes’ parameters (diameter, area, and volume) in B. bufo were significantly higher when compared with C. gariepinus and A. agama. The erythrocytes’ size was reported to have a direct association with chromosome number in fishes [19]. Increased erythrocytes’ size was noted with increasing chromosome number from diploid to triploid in C. gariepinus [20]. In vertebrates, erythrocytes’ size is significantly correlated with genome size [35, 36]. Among vertebrates, amphibians tend to have the largest erythrocytes size [37]. This was also observed in the current study. Amphibians were reported to have the largest genome size and the most significant variation in genome magnitude ranging from 0.95 pg in Platyplectrum ornatum to 140 pg in Necturus lewisi [38].
The large genome size in amphibians might cause a higher diameter, area and volume of erythrocytes in B. bufo compared with C. gariepinus and A. agama. Reptiles generally have a small genome size ranging from 1.1 pg to 5.4 pg [35]. Hence, the smaller erythrocytes size in A. agama compared with B. bufo. In fishes, erythrocyte sizes might differ in different species due to genome size. The erythrocytes of Australian lungfish with a genome magnitude of 50 pg were about 10-fold higher when compared with Siamese fighting fish with a genome size of 0.6 pg [35]. Therefore, genome size is essential in determining erythrocytes’ size in different vertebrates and even the same vertebrates in a different genus.
5. Conclusion
The current study indicated that C. gariepinus and B. bufo erythrocytes were better demonstrated with Giemsa stain while A. agama erythrocytes were better shown with H&E stain. This signifies that Erythrocytes’ affinity to stain varies in different vertebrates, probably due to environmental and genetic factors. The erythrocytes’ shape, size, area, and volume varies in different vertebrates, and the variation is associated with chromosome number and genome size.
Ethical Considerations
Compliance with ethical guidelines
The research was approved by the Department of Human Anatomy Ethical Committee (Code: UM/HA/UGP19.20-095) and conducted in accordance with National research council guide for the use of laboratory animals (8th edition).
Funding
This research did not receive any grant from funding agencies in the public, commercial, or non-profit sectors.
Authors' contributions
All authors equally contributed to preparing this article.
Conflict of interest
The authors declared no conflict of interest to declare.
References
- Arıkan H, Olgun K, Ilgaz Ç, Baran İ, Kumlutaş Y. Erythrocyte size and number in Neurergus strauchii (Urodela: Salamandridae). Russian Journal of Herpetology. 2003; 10(2):163-6. [Link]
- Hartenstein V. Blood cells and blood cell development in the animal kingdom. Annual Review of Cell and Developmental Biology. 2006; 22:677-12. [DOI:10.1146/annurev.cellbio.22.010605.093317] [PMID]
- Snyder GK, Sheafor BA. Red blood cells: Centerpiece in the evolution of the vertebrate circulatory system. American Zoologist. 1999; 39:189-98. [DOI:10.1093/icb/39.2.189]
- Frýdlová P, Hnízdo J, Chylíková L, Šimková O, Cikánová V, Velenský P, et al. Morphological characteristics of blood cells in monitor lizards: Is erythrocyte size linked to actual body size? Integretive Zoology. 2013; 8(s1):39-45. [DOI:10.1111/j.1749-4877.2012.00295.x] [PMID]
- Kumar MV. Morphometric studies of blood cells in Cyprinus Carpio, Ctenopharyngodan idella and Hypophthalmichthys Molitrix cultured fish in west Godavari region of Andhra Pradesh. International Journal of Fisheries and Aquatic Studies. 2016; 4:489-93. [Link]
- Wei J, Li Y, Wei L, Ding G, Fan X, Lin Z. Evolution of erythrocyte morphology in amphibians (Amphibia: Anura). Zoologia. 2015; 32:360-70. [DOI:10.1590/S1984-46702015000500005]
- GüL C, TosunoğLu M. Hematological reference intervals of four agamid lizard species from turkey (squamata: sauria: agamidae). Herpetozoa. 2011; 24(1-2):51-9. [Link]
- Cisneros GS, Thein SL. Recent advances in the treatment of sickle cell disease. Frontiers in Physiology. 2020; 11:435. [DOI:10.3389/fphys.2020.00435] [PMID] [PMCID]
- Svoboda O, Bartunek P. Origins of the vertebrate erythro/megakaryocytic system. BioMed Research International. 2015; 632171. [DOI:10.1155/2015/632171] [PMID] [PMCID]
- Alturkistani HA, Tashkandi FM, Mohammedsaleh ZM. Histological stains: A literature review and case study. Global Journal of Health Sciences. 2016; 8(3):72-9. [DOI:10.5539/gjhs.v8n3p72] [PMID] [PMCID]
- Dibal NI, Garba SH, Jacks TW. Morphological assessment of epididymal sperm in Wistar rats using different histological stains. Acta Veterinaria Eurasia. 2020; 46:132-6. [DOI:10.5152/actavet.2020.20018]
- Zhang Y, Haan K, Rivenson Y, Li J, Delis A, Ozcan A. Digital synthesis of histological stains using micro-structured and multiplexed virtual staining of label-free tissue. Light: Science and Applications. 2020; 9:1-13. [DOI:10.110.1109/JLT.2020.2986043]
- Rivenson Y, Wang H, Wei Z, Haan K, Zhang Y, Wu Y, et al. Virtual histological staining of unlabelled tissue-autofluorescence images via deep learning. Nature Biomedical Engineering. 2019; 3:466-77. [DOI:10.1038/s41551-019-0362-y] [PMID]
- Yamabayashi S. Periodic Acid-Schiff-Alcian Blue: A method for the differential staining of glycoproteins. The Histochemical Journal Volume. 1987; 19:565-71. [DOI:10.1007/BF01687364] [PMID]
- Isaiah IN, Florence O, Taiwo OJ, Augustina E, Terry On. Improvised eosin and leishman as morphological stain for sperm cell analysis; adult male wistar rats and rabbit as a model of study. Acta Scientific Microbiology. 2018; 1:18-20. [DOI:10.31080/ASMI.2018.01.0004]
- Barbalato L, Pillarisetty LS. Histology, red blood cell [Internet]. 2021 [Updated 2021 Nov 19]. Available from: [Link]
- Bhattacharjee D, Das S. Microscopic studies on erythrocytes of channa punctate exposed to commercial grade lindane. Brazilian Archives of Biology and Technology. 2017; 60:e17160341. [DOI:10.1590/1678-4324-2017160341]
- Shen Y, Wang D, Zhao J, Chen X. Fish red blood cells express immune genes and responses. Aquaculture and Fisheries. 2018; 3:14-21. [DOI:10.1016/j.aaf.2018.01.001]
- Zexia G, Weimin W, Yi Y, Abbas K, Dapeng L, Zou Guiwei Z, et al. Morphological studies of peripheral blood cells of the Chinese sturgeon, Acipenser sinensis. Fish Physiology and Biochemistry. 2007; 33:213-22. [DOI:10.1007/s10695-007-9133-x]
- Jalil N, Alim MA, Abol-Munafi AB, Ariffin NA, Waiho K, Sheriff SM. It is all in the blood: Erythrocyte characterization of triploid and diploid african catfish, clarias gariepinus. Journal of Fisheries and Aquatic Sciencei. 2016; 11(6):425-31. [DOI:10.3923/jfas.2016.425.431]
- Martins BO, Franco-Belussi L, Siqueira MS, Fernandes CE, Provete DB. The evolution of red blood cell shape in fishes. Journal of Evolutionary Biology. 2021; 34:537-48. [DOI:10.1111/jeb.13757] [PMID]
- Ponder E. Shape transformations in the red cells of camels, birds, reptiles, amphibia, and fishes. Journal of Experimental Biology. 1942; 19:215-9. [DOI:10.1242/jeb.19.3.215]
- Sato K, Uehara A, Kinoshita S, Nomura I, Yagi M, Tanizaki Y, et al. Flow cytometric analysis of Xenopus laevis and X. tropicalis blood cells using acridine orange. Scieitific Reports. 2018; 8:16245. [DOI:10.1038/s41598-018-34631-0] [PMID] [PMCID]
- Baehler RW, Catanxaro AJ, Stein JH, Hunter W. The Radiolabeled frog red blood cell. Circulation Research. 1973; 32(6):718-23. [DOI:10.1161/01.RES.32.6.718] [PMID]
- Kuramoto M. Relationships between number, size and shape of red blood cells in amphibians. Comparative Biochemistry and Physiology Part A: Physiology. 1981; 69:771-5. [DOI:10.1016/0300-9629(81)90170-5]
- Hota J, Das M, Mahapatra PK. Blood cell profile of the developing tadpoles and adults of the ornate frog, microhyla ornata (Anura: Microhylidae). International Journal of Zoology. 2013; 716183. [DOI:10.1155/2013/716183]
- Das M, Mahapatra PK. Blood cell profile of the Indian tree frog polypedates maculatus (gray, 1830), during larval development until metamorphosis (Anura: Rhacophoridae) Herpetozoa. 2015; 27:123 -35. [Link]
- Martınez-Silvestre A, Marco I, Rodriguez-Dominguez MA, Lavın S, Cuenca R. Morphology, cytochemical staining, and ultrastructural characteristics of the blood cells of the giant lizard of El Hierro (Gallotia simonyi). Research in Veterinary Science. 2005; 78(2):127-34. [DOI:10.1016/j.rvsc.2004.07.009] [PMID]
- Ponsen S, Talabmook C, Narkkong N, Aengwanich W. Blood cell characteristics and some hematological values of sand lizards (Leiolepis belliana rubritaeniata Martens 1961) in northeastern Thailand. International Journal of Zoological Research. 2008; 4:119-23. [DOI:10.3923/ijzr.2008.119.123]
- Sacchi R, Scali S, Cavirani V, Pupin F, Pellitteri-Rosa D, Zuffi MAL. Leukocyte differential counts and morphology from twelve European lizards. Italian Journal of Zoology. 2011; 78:418-26. [DOI:10.1080/11250003.2011.568014]
- Tsai C, Yu J, Wan Y, Fan P, Cheng T, Wang L. An alternative staining method for counting red-eared slider turtle (Trachemys scripta) blood cells using crystal violet in cells diluted with 0.45% sodium chloride. Journal of Veterinary Diagnostic Investigation. 2014; 26:610-5. [DOI:10.1177/1040638714542868] [PMID]
- Morera D, Roher N, Ribas L, Balasch JC, Do~nate C, Callol A, et al. RNASeq reveals an integrated immune response in nucleated erythrocytes. Plos One 2011; 6:e26998. [DOI:10.1371/journal.pone.0026998] [PMID] [PMCID]
- Finstad ØW, Dahle MK, Lindholm TH, Nyman IB, Løvoll M, Wallace C, et al. Piscine orthoreovirus (PRV) infects Atlantic salmon erythrocytes. Veterinary Research. 2014; 45(1):35. [DOI:10.1186/1297-9716-45-35] [PMID] [PMCID]
- Settembre C, Fraldi A, Medina DL, Ballabio A. Signals from the lysosome: A control centre for cellular clearance and energy metabolism. Nature Reviews. Molecular Cell Biology. 2013; 14:283-96. [DOI:10.1038/nrm3565] [PMID] [PMCID]
- Gregory TR. The bigger the c-value, the larger the cell: Genome size and red blood cell size in vertebrates. Blood Cells, Molecules, and Diseases. 2001; 27:830-43. [DOI:10.1006/bcmd.2001.0457] [PMID]
- Mueller RL, Gregory TR, Gregory SM, Hsieh A, Boore JL. Genome size, cell size, and the evolution of enucleated erythrocytes in attenuate salamanders. Zoology. 2008; 111(3):218-30. [DOI:10.1016/j.zool.2007.07.010] [PMID] [PMCID]
- Arikan H, Çiçek K. Haematology of amphibians and reptiles: A review. North Western Journal Zoology. 2014; 10(10):190-209. [Link]
- Liedtke HC, Gower DJ, Wilkinson M, Gomez-Mestre I. Macroevolutionary shift in the size of amphibian genomes and the role of life history and climate. Nature Ecology & Evolution. 2018; 2(11):1792-9. [DOI:10.1038/s41559-018-0674-4] [PMID]
Type of Study: Original |
Subject:
Histology
Received: 2021/11/6 | Accepted: 2022/04/6 | Published: 2022/02/26
Received: 2021/11/6 | Accepted: 2022/04/6 | Published: 2022/02/26
Send email to the article author
| Rights and permissions | |
 |
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License. |

Copyright © The Author(s);
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC-By-NC), which permits use, distribution, and reproduction in any medium, provided the original work is properly cited and is not used for commercial purposes.
Contact Information
