Sat, Jul 12, 2025
Volume 16, Issue 1 (Winter & Spring 2019)
ASJ 2019, 16(1): 65-80 |
Back to browse issues page

Download citation:
BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:



BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:
Dehghani Tafti E, Shojaei B, Tootian Z, Hashemnia S, Sheybani M T. Histological and Histochemical Study of Esophagus in Pheasant (Phasianus Colchicus) Embryo. ASJ 2019; 16 (1) :65-80
URL: http://anatomyjournal.ir/article-1-223-en.html
URL: http://anatomyjournal.ir/article-1-223-en.html
Elahe Dehghani Tafti1 

 , Bahador Shojaei2 , Zahra Tootian3 , Shadi Hashemnia2 , Mohammad Taghi Sheybani3
, Bahador Shojaei2 , Zahra Tootian3 , Shadi Hashemnia2 , Mohammad Taghi Sheybani3
, Bahador Shojaei2 , Zahra Tootian3 , Shadi Hashemnia2 , Mohammad Taghi Sheybani3
1- Phd student of Veterinary Anatomy and Embryology. Department of Basic Science, Faculty of Veterinary Medicine, University of Tehran, Tehran, Iran.
2- Department of Basic Sciences, Faculty of Veterinary Medicine, Shahid Bahonar University of Kerman, Kerman, Iran.
3- Department of Basic Sciences, Faculty of Veterinary Medicine, University of Tehran, Tehran, Iran.
2- Department of Basic Sciences, Faculty of Veterinary Medicine, Shahid Bahonar University of Kerman, Kerman, Iran.
3- Department of Basic Sciences, Faculty of Veterinary Medicine, University of Tehran, Tehran, Iran.
Full-Text [PDF 572 kb]
(2127 Downloads)
| Abstract (HTML) (5016 Views)


9th day of incubation
Two-layered cuboidal cells were developed and noticed more than ciliated pseudostratified columnar cells in cervical epithelium. However, the epithelial layer of thoracic esophagus was mostly composed of ciliated stratified cuboidal and rarely of simple cuboidal cells. In both parts of esophagus, the AB and PAS staining revealed acidic and neutral GACa, according to their blue and red colors, respectively.
10th day of incubation
The epithelial layer of cervical esophagus was composed of 2 simple and stratified cuboidal epithelium, with a large number of cilia. These cilia had covered the abdominal surface of the lumen, more than the dorsal side. A few capillaries were detected in the inner side of thoracic esophagus circular muscle of mesenchyme . The epithelium of the thoracic esophagus was similar to cervical part, but most of the cells were stratified cuboidal. The number of mucosal folds had increased to 10.
11th day of incubation
The development structures of layers of the cervical and thoracic esophagus did not differ from 10th day and were histologically similar.





12th day of incubation
The number of vessels in lamina propria increased. The outer longitudinal muscle was about to form the wall of thoracic esophagus. In the cervical part, the ciliated stratified cuboidal epithelium, gradually changed to the simple cuboidal from the bottom to the apex of folds. The epithelium of the thoracic part transferred into a 2-layered cuboidal epithelium (Figure 5).
13th day of incubation
In some regions of cervical part, the ciliated 2-layered cuboidal epithelium differentiated into stratified squamous. In the outermost layer of the mesenchyme, there was a cellular line, the progenitor of the outer longitudinal muscle. In this stage, mucosal muscle was visible in the thoracic part.
14th day of incubation
The epithelium of the cervical esophagus was mostly stratified squamous. The outermost muscle of cervical esophagus was about to generate. Ciliated squamous cells in some parts of the thoracic esophagus were recognized in this stage.


15th day of incubation
The epithelium of cervical esophagus was ciliated stratified squamous and stratified cuboidal in some parts.
In this stage, lamina propria and submucosa were about to separate by the formation of muscular layer of the mucosa. The epithelial tissue was mostly ciliated stratified squamous in the thoracic esophagus. In this part, the epithelial buds were arranged as circular groups of cells. Like the previous day, muscular layer of the mucosa was noticed continuously in folds of the thoracic part. The inner circular muscle was visible in layers. However, the outer longitudinal one was developing and did not form completely according to the Van Geison staining (Figure 6).
16th day of incubation
The structure of cervical and thoracic esophagus was similar to the previous day (Figure 7).
17th day of incubation
The epithelial layer of cervical esophagus was stratified squamous and thinner than the previous days. The muscular layer of the mucosa was developing almost continuously. However, the outer longitudinal muscle was observed as an incomplete layer. In this day, the gland progenitor cells were detected as buds on the base of folds of the thoracic esophagus. The number of epithelial buds was more than the previous day.
18th day of incubation
The epithelial buds appeared in the cervical esophagus. Outer longitudinal muscle was recognized as a continuous layer (Figure 8).
19th day of incubation
The mucosal layer of cervical esophagus was lined by stratified squamous cells with their cilia decreasing gradually. In some parts, epithelial buds were separated from the epithelium and seen independently in the lamina propria.
20th day of incubation
The epithelial tissue was stratified squamous and the number of epithelial buds increased. The structure of thoracic esophagus was similar to the previous day, but the epithelial buds were larger than the cervical one.
21st day of incubation
The epithelial layer of thoracic esophagus was stratified squamous with some vesicles in it (Figure 9). The cells of epithelial buds were organized in layers around the glands lumen. Vesicles were observed in the epithelium of thoracic part. Numerous large glands positively responded to PAS and Alcian Blue staining. According to the staining, the epithelial tissue also showed positive reaction to neutral and acid GAG’s (Figure 9).
22nd day of incubation
The lumen of cervical esophageal glands were observed and the cells of glands reacted to PAS and AB staining. In the thoracic part, the number of glands increased and the circular muscle layer was observed as continuous.
23rd day of incubation
Cervical and thoracic esophagus were same as the previous day.
24th day of incubation
PAS and AB staining showed the secretory activity of the epithelium, along the cervical and thoracic esophagus. Epithelial vesicles were visible, same as the 3 past days (Figure 10). The important point of these events are summarized in the Table 1.
4. Discussion
The esophageal development has been studied in different species. Among these studies, mammals have attracted more attention than other species. The initial study of embryogenesis of esophagus was performed on the epithelium of the pig esophagus, in 1905 [20]. The histogenesis of the human esophagus mucosa was studied in 1910 [21]. Development of esophagus epithelium has been discussed in 15th to 18th day of mouse embryonic period [22]. Also, the formation of different parts of esophagus has been investigated in prenatal period of the horse [23]. Moreover, esophagus development has been noted in the histogenetic studies of the embryonic gut of the other domestic mammals, like the camel and buffalo [24, 25].
Less attention has been paid on the morphogenesis of the esophagus in birds. Limited studies are available on the esophagus of chicken, turkey, pigeon, guinea fowl and partridge [8, 16, 17]. These studies discussed the sequence of layers formation, cell differentiation, changes of epithelial tissue and shape of lumen, separation of lamina propria from submucosa, appearance of glands and their secretory function during the incubation period. These studies showed variations in the time of appearance of the above-mentioned structures. This study was designed to explore esophagus histogenetic pattern of the pheasant. The results revealed that colchicus has many similarities to the partridge, as another member of phasianidae, in comparison to other bird families.
Ventura et al. studied the morphogenesis of Gallus gallus domesticus during 12th to the 20th day of incubation [3]. Gosomji performed the same investigation on the guinea fowl from 8th day of incubation onward [16]. Thus, the initial morphogenetic events of principal layer formation of the esophagus were disregarded by these researchers. Bellairs and Osmond studied the histogenesis of chick embryos from 50 h (stage 14) of incubation [5]. In the present study, in-line with the study by partridge [17], the lateral flexures were not complete until the end of the 4th day. Therefore, the morphogenetic events of the pheasant esophagus were detected from the 5th day. The rapid and daily histogenetic changes of the partridge esophagus [17], convinced us to collect the samples on each day of the incubation period.
Ventura studied the morphogenesis of Gallus gallus domesticus esophagus mucosa by PAS and AB staining [3]. However, Gosomji only used PAS staining for neutral GAG’s [16]. Also, in the evaluation of esophagus formation in emu bird and wild bird, these staining methods have been used for studying GAG’s and Van Geison was applied for detecting the appearance of collagen [13, 14]. Shehan et al. also used Van Geison in the histologic evaluation of geese esophagus [15]. In the present study, in addition to H&E, we used Van Geison to illustrate the formation of muscular layers, and PAS and AB staining to detect the neutral and acid GAG’s secretion in epithelial tissue and glands.
In the present study, esophagus was recognized as a compact cellular mass under the notochord and dorsal aorta, on the fifth day of incubation. This mass has been similarly observed on the fifth day in the partridge, but earlier, on 50th h of incubation, in the chick embryo [5]. The first time for appearance of esophageal has not been mentioned in the pigeon and turkey [8].
On the 5th day of incubation, both cervical and thoracic regions of esophagus were lined by pseudostratified columnar epithelium in the pheasant, partridge and fowl [8, 17]. This layer was reported as a simple columnar or seldomly pseudostratified in young embryos of chicken, turkey, guinea fowl and pigeon [8]. The epithelial cells of pheasant thoracic esophagus made a ciliated short pseudostratified columnar layer on the eighth day and differentiated into the cuboidal layer in both part of esophagus, similar to the fowl and partridge [8, 17].
In initial days of the second half of embryonic period, the esophagus epithelium of pheasant was going to change to stratified squamous, while the basal epithelial cells of partridge cervical esophagus has been reported as a stratified cuboidal layer on the last third of incubation period [17]. Ivey and Edgar observed 2-layered cuboidal epithelium in the chicken, turkey, guinea fowl and pigeon elder embryos that sometimes were transferred to squamous shape [8].
Gosomji et al. stated the gradual differentiation of epithelial surface to squamous on the last third of guinea fowl incubation period. They did not report any keratinization until the end of incubation period [16]. The differentiation of epithelium is associated with its function. Compensatory secretion activity of the epithelium is columnar in initial days of esophageal formation. Due to the protective role of mature form, the stratified squamous epithelium is visible at the end of incubation period. The continues secretion activity in the epithelium and the differentiation of the columnar epithelium into its mature form are important. Thus, the height of columnar cells reduced gradually in the middle of the incubation period and cuboidal cells were observed as transient cells in this period.
Due to the secretory function of the glands and accumulation of these secretions, some epithelial vesicles appear in continuation of the lumen of glands [8]. The epithelial vesicles of pheasant and partridge thoracic esophagus appeared on the 21st day of incubation. The same were appeared in the cervical esophagus of birds on the 24th embryonic day [17]. Other studies have not mentioned the appearance time of these vesicles.
Similar to the partridge [17], cilia are detected for the first time in pheasant esophagus in initial days of incubation period and exist until the last day of embryonic period. In the chicken and turkey, the cilia are observed up to the first 3-quarter and in the middle of the incubation period, respectively. In the guinea fowl, it is visible only at the end of the incubation period in the thoracic part and the pigeon embryo does not have cilia on its esophageal epithelium [8].
Apical cilia are recognized as essential cellular organelles in the regulation of developmental process. It has been stated that embryonic cells employ it to process incoming signals from morphogens and to generate and respond to mechanical signals [26]. Thus, in the initial days of incubation, the existence of cilia can be interpreted for its regulation role, but in the last days, it could have association with the nature of adult form of the esophagus in different species.
The basement membrane is a component of the extra matrix associated with the basal surface of adult and embryonic epithelia [4]. Basement membranes are required for the polarization of cells in both the embryo and adults. Cell polarity is an important initiating event in tissue development [27]. The appearance of basement membrane on the fifth day in the pheasant (present study) and partridge embryos [17] demonstrated the synchronization of endodermal differentiation into the epithelial tissue in these 2 species. The time of basement membrane formation is not mentioned in prior studies.
In molecular aspect, the interaction between epithelium and mesenchymal tissue is necessary for the gut development [1]. Therefore, it is expected that the esophageal mesenchyme become differentiated after its epithelium formation. The consequence of these cellular-molecular interactions can be detected by light microscope, as morphogenetic movements and sorting of mesenchymal cells. The sepration of pheasant trachea and esophagus mesenchyme was observed, 1 day after the differentiation of their epithelium into the thoracic region and , in the cervical region, 2 days later and, on the seventh day of incubation period. Consistent with our findings, prior research also reported the separation of partridge trachea and esophagus mesenchyme, one day after their epithelium differentiation on the sixth day of incubation [17].
The formation of muscular layers and muscular layer of the mucosa which shape esophageal wall, occur at different times in various species. The muscular layer enfolds the luminar structure of the esophagus as a coil and intensifies it. In pheasant and partridge [17], the cellular line for inner circular muscle formation is first detected in both parts of esophagus on the seventh day of incubation period. The outer longitudinal muscle was observed on the second half of incubation period in the pheasant thoracic esophagus and 2 days later in the cervical region. This is while its occurrence has been reported in both parts of partridge esophagus 1 day later than the pheasant thoracic region [17].
In the development of partridge (Tiramidae) esophagus, 3 rows of muscular layers were formed on the lamina propria, which consisted of inner longitudinal, middle circular and outer longitudinal layers [11]. Ivey discussed that the external longitudinal layer was either poorly developed or absent, and the submucosa was reduced greatly during the last quarter of incubation. Thus, the muscular layer of the mucosa appeared as a part of the muscular layer. The adult pigeon esophagus has been described without muscular layer of the mucosa but with an inner longitudinal and an outer circular layer [8].
The muscular layer of the mucosa is formed after muscular layer. The muscular layer of the mucosa increases firmness of the esophagus mucosa and regulates the dilation rate of the lumen. In this study, muscular layer of the mucosa appeared in thoracic esophagus in the middle of incubation period and 2 days later in the cervical region. In the partridge, this muscle appears 1 and 2 days later in the thoracic and cervical regions, respectively [17].
At the beginning of the last third of partridge incubation, the muscular layer of the mucosa is close to the muscular layer of the thoracic esophagus and they ae attached on the 19th day [17]. In pheasant, the muscular layer of the mucosa was extended into the mucosal folds on the second half of incubation period This extension was the beginning of formation of mucosal fold. Also, esophageal muscular layer of the mucosa of Japanese quail embryo extends into the epithelial folds [13]. However, this muscle extends into the folds of chicken, guinea fowl and emu esophagus, occasionally [8]. Considering the high rate of keratin concentration, Van Gieson’s stain demarcated the mature form of muscle in the second half of the incubation period, in the current investigation.
The development of glands begins from the epithelial cells bud, which agglomerates the connective tissue. These cells recede gradually from the surface epithelium and excavate in the mesenchyme. The circular clusters of cells form primordia of esophageal glands [3]. The pheasant and partridge [17] epithelial buds of thoracic esophagus were noticed at the last third of incubation period. Howver, they were detected in the cervical region of partridge esophagus [17], a day later than pheasant. These buds were visible in thoracic esophagus of chicken embryos and guinea fowls in the middle third of incubation period [8, 17]. Gosomji reported that the epithelial cells began to proliferate at their basal pole to form clusters of primordial esophageal glands at the last third of guinea fowl incubation period [16].
At the beginning of the development, the mucosal layer of the esophagus has secretory epithelial cells that secrete neutral and acid GAG’s. The secretory activity of these cells is probably meant to compensate the lack of mucosa glands [3]. During the glands formation and at the beginning of their secretion, the height of epithelial cells and their compensate secretion activity reduced gradually in the present study, which is same as the partridge [17]. The acidic materials were detected in the lumen of esophagus, in initial days. Considering that these secretions were not observed in the following days, they could not be as a result of compensate activity of epithelial tissue. According to the study by Ivey, this material might be yolk from the yolk sac or a mixture of yolk and albumen [8].
In the pheasant and partridge [17], compensate activity of both parts of esophagus epithelium begin in the initial days of second half of incubation period. However positive reaction pf GAG’s to PAS staining was observed later in partridge esophagus [17]. In the Gallus gallus domesticus, the esophageal epithelial cells are positive for the production of neutral and acid GAG’s, in the second half of the incubation period [3]. The secretion activity of epithelium in both parts of pheasant esophagus continued until the end of incubation period.
At the end of embryonic period, the lumen of glands appears by arrangement of the epithelial buds cells [3]. Similar to the study on partridge [17], the lumen of glands were visible in cervical and thoracic regions of pheasant esophagus near the end of incubation period. In the turkey, the lumen of mucos glands were only detected in the cervical part, near the end of the incubation period [8]. A wide lumen is created in each of the primordial oesophageal glands displacing the cells from the center to the periphery in the middle and at the end of embryonic period of the Gallus gallus domesticus and guinea fowl embryos, respectively [3, 16].
The neutral and acid GAG’s produce slightly and high viscous mucus, respectively. The neutral GAG’s favors the digestive transit, lubricates food particles, allows them to be pushed to the next segment. The acid GAG’s are very important to trap particles and form a protective barrier against mechanical friction and the action of the digestive acid [3]. The beginning of the secretions which had positive reaction to AB and PAS staining was noted in the pheasant thoracic esophagus on the 21st day of incubation period and a day later in the cervical region. These observations are reported 3 days earlier and 2 days later in the thoracic and cervical regions of partridge esophagus, respectively [17].
The primordial glandular tissue of Gallus gallus domesticus was weakly positive to AB and PAS staining in the last quarter of embryonic period. However, strong positive reaction to these staining in the next days showed full production of neutral and acid GAG’s in these species [3]. In the guinea fowl, near the end of the incubation period, the epithelium and primordial glands demonstrated weak reactions to PAS. However, the glands show mucosal characteristics at the end of incubation period [16].
In conclusion, the this study declared that the most esophageal developmental events of the pheasant occur in the first 2-thirds of the incubation period. Most of these events were observed in the thoracic esophagus earlier than cervical region. Comparison of our results with similar studieson the avian esophagus development, indicates that the sequence of the developmental events in pheasant are similar to the other birds; however, the three are variations in onset time of events. The differentiation of the epithelial cells into stratified squamous occur in the initial days of second half of pheasant incubation period., but However, it is seen at the last third stage in partridge and guinea fowl.
Cilia appear in the initial day of pheasant and partridge embryonic period. However, previous studies reported its occurrence almost in the middle or near the end of incubation period. The esophageal glands of pheasant and partridge form at the last third of incubation time, but they appear almost in the second half of chicken and middle third of turkey embryonic period. According to this study and the other investigations, the formation of lumen of glands is obsereved near the end of incubation. However, it occurs in the middle of the embryonic period among Gallus gallus domesticus.
Some of these variations can be related to the differences in the length of the incubation period in different species. In this case, a correlation is expected between the variations of developmental events and the length of incubation periods. The present study indicates that this prospect works for pheasant and partridge as 2 members of Phasianidae. However, this rule is not reported regarding other studied species.
Ethical Considerations
Compliance with ethical guidelines
All steps of this study were approved by the Animal Ethics Committee, based on the Research Council of the Veterinary Faculty of Tehran University.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Authors contributions
All authors have read and approved the manuscript.
Conflict of interest
The authors declared no conflict of interest.
Acknowledgments
This research was financially supported by the Research Council of Faculty of Veterinary Medicine, University of Tehran.
References
Full-Text: (4057 Views)
1. Introduction
Sequence The developmental model of organs are presented by histogenesis and its sequences. These models provide basic information for studying factors involved in differentiation of embryonic cells to specific structures. Also, these models are necessary for researchers to evaluate the influence of different agents during the critical period of organ formation [1, 2]. The birds are investigated as models in experimental embryologic studies, more than other animals. Accessibility, incubation period and economic viability made these embryo models prominent [3]. The normal embryology of avian organs is explored in many studies and essential for researchers, [1, 4, 5].
The oesophagus of birds reflects histological differences between those and mammals including glandular lamina propria [6, 7], different extensions of muscularis mucosae into the mucosal folds [8-10], and absence of muscularis mucosa in some birds [8, 11]. Consequently, these differences result from different sequential events of the esophagus histogenesis in these 2 classes of animals. Moreover, studies on the morphology and histology of esophagus of some birds [11-15] demonstrated differences in the shape and structure of this organ. The esophagus originates from the foregut in all species. Thus, such differences can be described according to variations in the histogenetic events during organ formation.
Esophagus morphogenesis has been studied in domestic fowl, turkey, guinea fowl, pigeon and partridge [3, 8, 16, 17]. These studies claimed that the initial scaffold of the esophagus is made from anterior part of the gut as a layer of endodermal tube [17, 18]. They also stated that the mesenchyme, surrounding the endodermal tube differentiates into the connective tissue and muscular layers, gradually [16]. However, the time of layers formation, separation of lamina propria and submucosa, appearance of the glands and their secretory function, lumen shape [3] and the differentiation of epithelial tissue from columnar into stratified squamous show some differences [8]. This heterogeneity, may be due to differences in their nutritional behavior, length of incubation period and/or evolutionary origin of avian species.
As the studied birds belong to different families, the mentioned histogenetic variations may be discussable according to species specific characters. The present study was designed to examine the presence of these differences in the members of a family. Pheasant (colchicus) can be studied as an embryological model for experiments. It belongs to Phasianidae which one of its members, chukar partridge, has recently been mentioned in esophagus development. The present work is the first to describe, the histomorphology of the esophagus in this bird to produce a developmental events timetable of this organ. Moreover, the results of this study can explain the resemblance between the developmental pattern of pheasant esophagus and the chukar partridge.
2. Material and Methods
A total of 250 embryonated pheasant (colchicus) eggs were incubated at 37.7oC with humidity of 65%. At the end of incubation period, the temperature and humidity were changed respectively to 36.7oC and 75% for hatching eggs. From the fourth day of incubation, at least 4 alive embryos were euthanized and collected, then fixed in 10℅ buffered formalin solution, each day. In order to have better fixation in the large embryos (11th-24th days), the neck and body were dissected and fixed. Then paraffin blocks were prepared from fixed samples and the 5 µm sections were obtained using a MR2258 microtome (Histo-Line, Pantigliate, Italy). The slides were stained with Hematoxylin and Eosin (H&E), Periodic Acid-Schiff (PAS) and Alcian Blue (AB), Van Gieson (VG) (Merck, KGaA, 64271, Darmstadt, Germany) methods [19] and studied by a light microscope.
Initially, the sections were deparaffinized and hydrated to water. For Periodic Acid-Schiff staining, the samples were oxidized in periodic acid solution 0.5 gm for 5 minutes and rinsed in distilled water. They were placed in Schiff reagent for 15 minutes. Then, they were changed in sodium bisulfite, 3 times, every 2 minutes and washed in running tap water for 5 minutes. They were counterstained in Harris’ hematoxylin for 1 minute and washed again in running water for 5 minutes. Eventually, the samples were dehydrated and coverslipped using a synthetic mounting medium [19].
In Alcian BlueVan Gieson protocol, the sections were stained in weigert’s hematoxylin solution for 20 minutes and then washed in running water for 5 minutes. After that, samples were placed in acid alcohol for 10 to 20 seconds and washed in running water. Then they were stained in Alcian blue solution for 10 minutes. After being washed in tap water, samples were stained in Van Gieson’s solution for 5 to 10 minutes. Finally, they were dehydrated through 95% alcohol and 2 changes of absolute alcohol, 3 minutes each. Then samples were cleared in xylene and mounted with resinous mounting medium [19].
The general structure of the esophagus tissue and the appearance of different layers and cells were examined by H&E staining. The neutral and acid Glycosaminoglycans (GAGs) secretions were studied with PAS and AB staining. Eventually, the VG staining helped to distinguish the connective tissue via the red collagen fibers. Muscles were also distinguished according to their color.
3. Results
4th day of incubation
The esophageal primordial was not observed, due to incomplete folding of the embryonic disc.
5th day of incubation
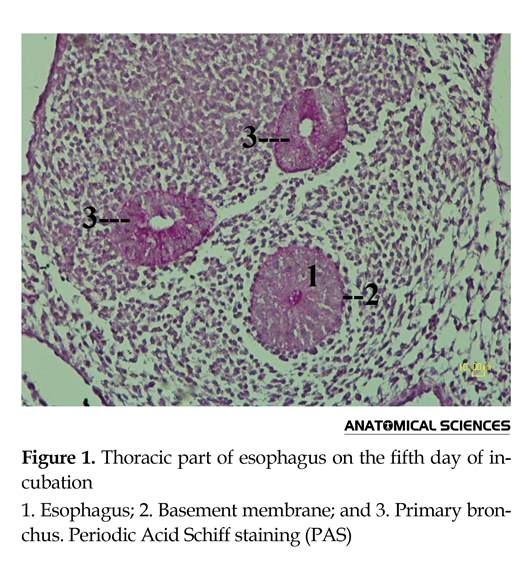
The esophagus was observed as a compact mass under the notochord and the branches of aortae and above the trachea. The lumen of esophagus was small and covered by pseudo-stratified columnar epithelium. It was filled with acidophilic materials. A homogenous mesenchymal connective tissue was visible around the epithelia of the esophagus and trachea. According to the PAS staining, the basal lamina was observed as a red ring surrounding the epithelial tissue (Figure 1).
6th day of incubation
The cervical esophagus lumen was decreased. However, the thoracic esophagus was same as the previous day. The mucosa was lined by the pseudostratified columnar epithelium, with some cilia growing. The mesenchymal boundaries of trachea and esophagus were about to be distinguished in the thoracic part.
7th day of incubation
The cervical esophagus lumen was completely closed. However, it was open in the thoracic region and covered by pseudostratified columnar epithelium. In the thoracic part, the mesenchymal boundary of the esophagus and trachea was completely distinct , but in the cervical part, it was just about to be distinguished from one another. In this stage, the progenitor mesenchymal cells of inner circular muscle began to differentiate in both parts of esophagus, for the first time. Also, we observed a mesenchymal line out of the epithelial tissue, which was generating lamina propria (Figure 2 and 3).
8th day of incubation
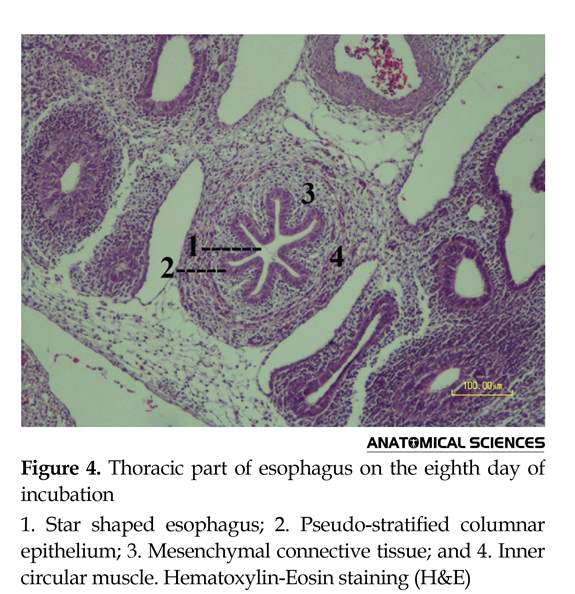
The epithelial layer of cervical esophagus was mostly composed of pseudostratified ciliated columnar cells. There were a number of cilia on abdominal side of the lumen, which were more visible on this side, compared to the dorsal side. Some folds were observed in cervical esophagus. Although the epithelial layer was stratified in some parts of the thoracic part, the height of epithelial cells was reduced and mostly changed to the short ciliated pseudostratified columnar. Six folds formed the mucosal layer, and changed the appearance of lumen to a star shape. The underlying compact mesenchymal cells entered into the epithelial folds to increase their thickness. The looser mesenchymal cells were observed between the compact area and circular muscle. In this stage, progenitors of smooth muscle cells were observed in a spindle shape and relative vessels have reached to their vicinity (Figure 4).
Sequence The developmental model of organs are presented by histogenesis and its sequences. These models provide basic information for studying factors involved in differentiation of embryonic cells to specific structures. Also, these models are necessary for researchers to evaluate the influence of different agents during the critical period of organ formation [1, 2]. The birds are investigated as models in experimental embryologic studies, more than other animals. Accessibility, incubation period and economic viability made these embryo models prominent [3]. The normal embryology of avian organs is explored in many studies and essential for researchers, [1, 4, 5].
The oesophagus of birds reflects histological differences between those and mammals including glandular lamina propria [6, 7], different extensions of muscularis mucosae into the mucosal folds [8-10], and absence of muscularis mucosa in some birds [8, 11]. Consequently, these differences result from different sequential events of the esophagus histogenesis in these 2 classes of animals. Moreover, studies on the morphology and histology of esophagus of some birds [11-15] demonstrated differences in the shape and structure of this organ. The esophagus originates from the foregut in all species. Thus, such differences can be described according to variations in the histogenetic events during organ formation.
Esophagus morphogenesis has been studied in domestic fowl, turkey, guinea fowl, pigeon and partridge [3, 8, 16, 17]. These studies claimed that the initial scaffold of the esophagus is made from anterior part of the gut as a layer of endodermal tube [17, 18]. They also stated that the mesenchyme, surrounding the endodermal tube differentiates into the connective tissue and muscular layers, gradually [16]. However, the time of layers formation, separation of lamina propria and submucosa, appearance of the glands and their secretory function, lumen shape [3] and the differentiation of epithelial tissue from columnar into stratified squamous show some differences [8]. This heterogeneity, may be due to differences in their nutritional behavior, length of incubation period and/or evolutionary origin of avian species.
As the studied birds belong to different families, the mentioned histogenetic variations may be discussable according to species specific characters. The present study was designed to examine the presence of these differences in the members of a family. Pheasant (colchicus) can be studied as an embryological model for experiments. It belongs to Phasianidae which one of its members, chukar partridge, has recently been mentioned in esophagus development. The present work is the first to describe, the histomorphology of the esophagus in this bird to produce a developmental events timetable of this organ. Moreover, the results of this study can explain the resemblance between the developmental pattern of pheasant esophagus and the chukar partridge.
2. Material and Methods
A total of 250 embryonated pheasant (colchicus) eggs were incubated at 37.7oC with humidity of 65%. At the end of incubation period, the temperature and humidity were changed respectively to 36.7oC and 75% for hatching eggs. From the fourth day of incubation, at least 4 alive embryos were euthanized and collected, then fixed in 10℅ buffered formalin solution, each day. In order to have better fixation in the large embryos (11th-24th days), the neck and body were dissected and fixed. Then paraffin blocks were prepared from fixed samples and the 5 µm sections were obtained using a MR2258 microtome (Histo-Line, Pantigliate, Italy). The slides were stained with Hematoxylin and Eosin (H&E), Periodic Acid-Schiff (PAS) and Alcian Blue (AB), Van Gieson (VG) (Merck, KGaA, 64271, Darmstadt, Germany) methods [19] and studied by a light microscope.
Initially, the sections were deparaffinized and hydrated to water. For Periodic Acid-Schiff staining, the samples were oxidized in periodic acid solution 0.5 gm for 5 minutes and rinsed in distilled water. They were placed in Schiff reagent for 15 minutes. Then, they were changed in sodium bisulfite, 3 times, every 2 minutes and washed in running tap water for 5 minutes. They were counterstained in Harris’ hematoxylin for 1 minute and washed again in running water for 5 minutes. Eventually, the samples were dehydrated and coverslipped using a synthetic mounting medium [19].
In Alcian BlueVan Gieson protocol, the sections were stained in weigert’s hematoxylin solution for 20 minutes and then washed in running water for 5 minutes. After that, samples were placed in acid alcohol for 10 to 20 seconds and washed in running water. Then they were stained in Alcian blue solution for 10 minutes. After being washed in tap water, samples were stained in Van Gieson’s solution for 5 to 10 minutes. Finally, they were dehydrated through 95% alcohol and 2 changes of absolute alcohol, 3 minutes each. Then samples were cleared in xylene and mounted with resinous mounting medium [19].
The general structure of the esophagus tissue and the appearance of different layers and cells were examined by H&E staining. The neutral and acid Glycosaminoglycans (GAGs) secretions were studied with PAS and AB staining. Eventually, the VG staining helped to distinguish the connective tissue via the red collagen fibers. Muscles were also distinguished according to their color.
3. Results
4th day of incubation
The esophageal primordial was not observed, due to incomplete folding of the embryonic disc.
5th day of incubation
The esophagus was observed as a compact mass under the notochord and the branches of aortae and above the trachea. The lumen of esophagus was small and covered by pseudo-stratified columnar epithelium. It was filled with acidophilic materials. A homogenous mesenchymal connective tissue was visible around the epithelia of the esophagus and trachea. According to the PAS staining, the basal lamina was observed as a red ring surrounding the epithelial tissue (Figure 1).
6th day of incubation
The cervical esophagus lumen was decreased. However, the thoracic esophagus was same as the previous day. The mucosa was lined by the pseudostratified columnar epithelium, with some cilia growing. The mesenchymal boundaries of trachea and esophagus were about to be distinguished in the thoracic part.
7th day of incubation
The cervical esophagus lumen was completely closed. However, it was open in the thoracic region and covered by pseudostratified columnar epithelium. In the thoracic part, the mesenchymal boundary of the esophagus and trachea was completely distinct , but in the cervical part, it was just about to be distinguished from one another. In this stage, the progenitor mesenchymal cells of inner circular muscle began to differentiate in both parts of esophagus, for the first time. Also, we observed a mesenchymal line out of the epithelial tissue, which was generating lamina propria (Figure 2 and 3).
8th day of incubation
The epithelial layer of cervical esophagus was mostly composed of pseudostratified ciliated columnar cells. There were a number of cilia on abdominal side of the lumen, which were more visible on this side, compared to the dorsal side. Some folds were observed in cervical esophagus. Although the epithelial layer was stratified in some parts of the thoracic part, the height of epithelial cells was reduced and mostly changed to the short ciliated pseudostratified columnar. Six folds formed the mucosal layer, and changed the appearance of lumen to a star shape. The underlying compact mesenchymal cells entered into the epithelial folds to increase their thickness. The looser mesenchymal cells were observed between the compact area and circular muscle. In this stage, progenitors of smooth muscle cells were observed in a spindle shape and relative vessels have reached to their vicinity (Figure 4).
9th day of incubation
Two-layered cuboidal cells were developed and noticed more than ciliated pseudostratified columnar cells in cervical epithelium. However, the epithelial layer of thoracic esophagus was mostly composed of ciliated stratified cuboidal and rarely of simple cuboidal cells. In both parts of esophagus, the AB and PAS staining revealed acidic and neutral GACa, according to their blue and red colors, respectively.
10th day of incubation
The epithelial layer of cervical esophagus was composed of 2 simple and stratified cuboidal epithelium, with a large number of cilia. These cilia had covered the abdominal surface of the lumen, more than the dorsal side. A few capillaries were detected in the inner side of thoracic esophagus circular muscle of mesenchyme . The epithelium of the thoracic esophagus was similar to cervical part, but most of the cells were stratified cuboidal. The number of mucosal folds had increased to 10.
11th day of incubation
The development structures of layers of the cervical and thoracic esophagus did not differ from 10th day and were histologically similar.
12th day of incubation
The number of vessels in lamina propria increased. The outer longitudinal muscle was about to form the wall of thoracic esophagus. In the cervical part, the ciliated stratified cuboidal epithelium, gradually changed to the simple cuboidal from the bottom to the apex of folds. The epithelium of the thoracic part transferred into a 2-layered cuboidal epithelium (Figure 5).
13th day of incubation
In some regions of cervical part, the ciliated 2-layered cuboidal epithelium differentiated into stratified squamous. In the outermost layer of the mesenchyme, there was a cellular line, the progenitor of the outer longitudinal muscle. In this stage, mucosal muscle was visible in the thoracic part.
14th day of incubation
The epithelium of the cervical esophagus was mostly stratified squamous. The outermost muscle of cervical esophagus was about to generate. Ciliated squamous cells in some parts of the thoracic esophagus were recognized in this stage.
15th day of incubation
The epithelium of cervical esophagus was ciliated stratified squamous and stratified cuboidal in some parts.
In this stage, lamina propria and submucosa were about to separate by the formation of muscular layer of the mucosa. The epithelial tissue was mostly ciliated stratified squamous in the thoracic esophagus. In this part, the epithelial buds were arranged as circular groups of cells. Like the previous day, muscular layer of the mucosa was noticed continuously in folds of the thoracic part. The inner circular muscle was visible in layers. However, the outer longitudinal one was developing and did not form completely according to the Van Geison staining (Figure 6).
16th day of incubation
The structure of cervical and thoracic esophagus was similar to the previous day (Figure 7).
17th day of incubation
The epithelial layer of cervical esophagus was stratified squamous and thinner than the previous days. The muscular layer of the mucosa was developing almost continuously. However, the outer longitudinal muscle was observed as an incomplete layer. In this day, the gland progenitor cells were detected as buds on the base of folds of the thoracic esophagus. The number of epithelial buds was more than the previous day.
18th day of incubation
The epithelial buds appeared in the cervical esophagus. Outer longitudinal muscle was recognized as a continuous layer (Figure 8).
19th day of incubation
The mucosal layer of cervical esophagus was lined by stratified squamous cells with their cilia decreasing gradually. In some parts, epithelial buds were separated from the epithelium and seen independently in the lamina propria.
20th day of incubation
The epithelial tissue was stratified squamous and the number of epithelial buds increased. The structure of thoracic esophagus was similar to the previous day, but the epithelial buds were larger than the cervical one.
21st day of incubation
The epithelial layer of thoracic esophagus was stratified squamous with some vesicles in it (Figure 9). The cells of epithelial buds were organized in layers around the glands lumen. Vesicles were observed in the epithelium of thoracic part. Numerous large glands positively responded to PAS and Alcian Blue staining. According to the staining, the epithelial tissue also showed positive reaction to neutral and acid GAG’s (Figure 9).
22nd day of incubation
The lumen of cervical esophageal glands were observed and the cells of glands reacted to PAS and AB staining. In the thoracic part, the number of glands increased and the circular muscle layer was observed as continuous.
23rd day of incubation
Cervical and thoracic esophagus were same as the previous day.
24th day of incubation
PAS and AB staining showed the secretory activity of the epithelium, along the cervical and thoracic esophagus. Epithelial vesicles were visible, same as the 3 past days (Figure 10). The important point of these events are summarized in the Table 1.
4. Discussion
The esophageal development has been studied in different species. Among these studies, mammals have attracted more attention than other species. The initial study of embryogenesis of esophagus was performed on the epithelium of the pig esophagus, in 1905 [20]. The histogenesis of the human esophagus mucosa was studied in 1910 [21]. Development of esophagus epithelium has been discussed in 15th to 18th day of mouse embryonic period [22]. Also, the formation of different parts of esophagus has been investigated in prenatal period of the horse [23]. Moreover, esophagus development has been noted in the histogenetic studies of the embryonic gut of the other domestic mammals, like the camel and buffalo [24, 25].
Less attention has been paid on the morphogenesis of the esophagus in birds. Limited studies are available on the esophagus of chicken, turkey, pigeon, guinea fowl and partridge [8, 16, 17]. These studies discussed the sequence of layers formation, cell differentiation, changes of epithelial tissue and shape of lumen, separation of lamina propria from submucosa, appearance of glands and their secretory function during the incubation period. These studies showed variations in the time of appearance of the above-mentioned structures. This study was designed to explore esophagus histogenetic pattern of the pheasant. The results revealed that colchicus has many similarities to the partridge, as another member of phasianidae, in comparison to other bird families.
Ventura et al. studied the morphogenesis of Gallus gallus domesticus during 12th to the 20th day of incubation [3]. Gosomji performed the same investigation on the guinea fowl from 8th day of incubation onward [16]. Thus, the initial morphogenetic events of principal layer formation of the esophagus were disregarded by these researchers. Bellairs and Osmond studied the histogenesis of chick embryos from 50 h (stage 14) of incubation [5]. In the present study, in-line with the study by partridge [17], the lateral flexures were not complete until the end of the 4th day. Therefore, the morphogenetic events of the pheasant esophagus were detected from the 5th day. The rapid and daily histogenetic changes of the partridge esophagus [17], convinced us to collect the samples on each day of the incubation period.
Ventura studied the morphogenesis of Gallus gallus domesticus esophagus mucosa by PAS and AB staining [3]. However, Gosomji only used PAS staining for neutral GAG’s [16]. Also, in the evaluation of esophagus formation in emu bird and wild bird, these staining methods have been used for studying GAG’s and Van Geison was applied for detecting the appearance of collagen [13, 14]. Shehan et al. also used Van Geison in the histologic evaluation of geese esophagus [15]. In the present study, in addition to H&E, we used Van Geison to illustrate the formation of muscular layers, and PAS and AB staining to detect the neutral and acid GAG’s secretion in epithelial tissue and glands.
In the present study, esophagus was recognized as a compact cellular mass under the notochord and dorsal aorta, on the fifth day of incubation. This mass has been similarly observed on the fifth day in the partridge, but earlier, on 50th h of incubation, in the chick embryo [5]. The first time for appearance of esophageal has not been mentioned in the pigeon and turkey [8].
On the 5th day of incubation, both cervical and thoracic regions of esophagus were lined by pseudostratified columnar epithelium in the pheasant, partridge and fowl [8, 17]. This layer was reported as a simple columnar or seldomly pseudostratified in young embryos of chicken, turkey, guinea fowl and pigeon [8]. The epithelial cells of pheasant thoracic esophagus made a ciliated short pseudostratified columnar layer on the eighth day and differentiated into the cuboidal layer in both part of esophagus, similar to the fowl and partridge [8, 17].
In initial days of the second half of embryonic period, the esophagus epithelium of pheasant was going to change to stratified squamous, while the basal epithelial cells of partridge cervical esophagus has been reported as a stratified cuboidal layer on the last third of incubation period [17]. Ivey and Edgar observed 2-layered cuboidal epithelium in the chicken, turkey, guinea fowl and pigeon elder embryos that sometimes were transferred to squamous shape [8].
Gosomji et al. stated the gradual differentiation of epithelial surface to squamous on the last third of guinea fowl incubation period. They did not report any keratinization until the end of incubation period [16]. The differentiation of epithelium is associated with its function. Compensatory secretion activity of the epithelium is columnar in initial days of esophageal formation. Due to the protective role of mature form, the stratified squamous epithelium is visible at the end of incubation period. The continues secretion activity in the epithelium and the differentiation of the columnar epithelium into its mature form are important. Thus, the height of columnar cells reduced gradually in the middle of the incubation period and cuboidal cells were observed as transient cells in this period.
Due to the secretory function of the glands and accumulation of these secretions, some epithelial vesicles appear in continuation of the lumen of glands [8]. The epithelial vesicles of pheasant and partridge thoracic esophagus appeared on the 21st day of incubation. The same were appeared in the cervical esophagus of birds on the 24th embryonic day [17]. Other studies have not mentioned the appearance time of these vesicles.
Similar to the partridge [17], cilia are detected for the first time in pheasant esophagus in initial days of incubation period and exist until the last day of embryonic period. In the chicken and turkey, the cilia are observed up to the first 3-quarter and in the middle of the incubation period, respectively. In the guinea fowl, it is visible only at the end of the incubation period in the thoracic part and the pigeon embryo does not have cilia on its esophageal epithelium [8].
Apical cilia are recognized as essential cellular organelles in the regulation of developmental process. It has been stated that embryonic cells employ it to process incoming signals from morphogens and to generate and respond to mechanical signals [26]. Thus, in the initial days of incubation, the existence of cilia can be interpreted for its regulation role, but in the last days, it could have association with the nature of adult form of the esophagus in different species.
The basement membrane is a component of the extra matrix associated with the basal surface of adult and embryonic epithelia [4]. Basement membranes are required for the polarization of cells in both the embryo and adults. Cell polarity is an important initiating event in tissue development [27]. The appearance of basement membrane on the fifth day in the pheasant (present study) and partridge embryos [17] demonstrated the synchronization of endodermal differentiation into the epithelial tissue in these 2 species. The time of basement membrane formation is not mentioned in prior studies.
In molecular aspect, the interaction between epithelium and mesenchymal tissue is necessary for the gut development [1]. Therefore, it is expected that the esophageal mesenchyme become differentiated after its epithelium formation. The consequence of these cellular-molecular interactions can be detected by light microscope, as morphogenetic movements and sorting of mesenchymal cells. The sepration of pheasant trachea and esophagus mesenchyme was observed, 1 day after the differentiation of their epithelium into the thoracic region and , in the cervical region, 2 days later and, on the seventh day of incubation period. Consistent with our findings, prior research also reported the separation of partridge trachea and esophagus mesenchyme, one day after their epithelium differentiation on the sixth day of incubation [17].
The formation of muscular layers and muscular layer of the mucosa which shape esophageal wall, occur at different times in various species. The muscular layer enfolds the luminar structure of the esophagus as a coil and intensifies it. In pheasant and partridge [17], the cellular line for inner circular muscle formation is first detected in both parts of esophagus on the seventh day of incubation period. The outer longitudinal muscle was observed on the second half of incubation period in the pheasant thoracic esophagus and 2 days later in the cervical region. This is while its occurrence has been reported in both parts of partridge esophagus 1 day later than the pheasant thoracic region [17].
In the development of partridge (Tiramidae) esophagus, 3 rows of muscular layers were formed on the lamina propria, which consisted of inner longitudinal, middle circular and outer longitudinal layers [11]. Ivey discussed that the external longitudinal layer was either poorly developed or absent, and the submucosa was reduced greatly during the last quarter of incubation. Thus, the muscular layer of the mucosa appeared as a part of the muscular layer. The adult pigeon esophagus has been described without muscular layer of the mucosa but with an inner longitudinal and an outer circular layer [8].
The muscular layer of the mucosa is formed after muscular layer. The muscular layer of the mucosa increases firmness of the esophagus mucosa and regulates the dilation rate of the lumen. In this study, muscular layer of the mucosa appeared in thoracic esophagus in the middle of incubation period and 2 days later in the cervical region. In the partridge, this muscle appears 1 and 2 days later in the thoracic and cervical regions, respectively [17].
At the beginning of the last third of partridge incubation, the muscular layer of the mucosa is close to the muscular layer of the thoracic esophagus and they ae attached on the 19th day [17]. In pheasant, the muscular layer of the mucosa was extended into the mucosal folds on the second half of incubation period This extension was the beginning of formation of mucosal fold. Also, esophageal muscular layer of the mucosa of Japanese quail embryo extends into the epithelial folds [13]. However, this muscle extends into the folds of chicken, guinea fowl and emu esophagus, occasionally [8]. Considering the high rate of keratin concentration, Van Gieson’s stain demarcated the mature form of muscle in the second half of the incubation period, in the current investigation.
The development of glands begins from the epithelial cells bud, which agglomerates the connective tissue. These cells recede gradually from the surface epithelium and excavate in the mesenchyme. The circular clusters of cells form primordia of esophageal glands [3]. The pheasant and partridge [17] epithelial buds of thoracic esophagus were noticed at the last third of incubation period. Howver, they were detected in the cervical region of partridge esophagus [17], a day later than pheasant. These buds were visible in thoracic esophagus of chicken embryos and guinea fowls in the middle third of incubation period [8, 17]. Gosomji reported that the epithelial cells began to proliferate at their basal pole to form clusters of primordial esophageal glands at the last third of guinea fowl incubation period [16].
At the beginning of the development, the mucosal layer of the esophagus has secretory epithelial cells that secrete neutral and acid GAG’s. The secretory activity of these cells is probably meant to compensate the lack of mucosa glands [3]. During the glands formation and at the beginning of their secretion, the height of epithelial cells and their compensate secretion activity reduced gradually in the present study, which is same as the partridge [17]. The acidic materials were detected in the lumen of esophagus, in initial days. Considering that these secretions were not observed in the following days, they could not be as a result of compensate activity of epithelial tissue. According to the study by Ivey, this material might be yolk from the yolk sac or a mixture of yolk and albumen [8].
In the pheasant and partridge [17], compensate activity of both parts of esophagus epithelium begin in the initial days of second half of incubation period. However positive reaction pf GAG’s to PAS staining was observed later in partridge esophagus [17]. In the Gallus gallus domesticus, the esophageal epithelial cells are positive for the production of neutral and acid GAG’s, in the second half of the incubation period [3]. The secretion activity of epithelium in both parts of pheasant esophagus continued until the end of incubation period.
At the end of embryonic period, the lumen of glands appears by arrangement of the epithelial buds cells [3]. Similar to the study on partridge [17], the lumen of glands were visible in cervical and thoracic regions of pheasant esophagus near the end of incubation period. In the turkey, the lumen of mucos glands were only detected in the cervical part, near the end of the incubation period [8]. A wide lumen is created in each of the primordial oesophageal glands displacing the cells from the center to the periphery in the middle and at the end of embryonic period of the Gallus gallus domesticus and guinea fowl embryos, respectively [3, 16].
The neutral and acid GAG’s produce slightly and high viscous mucus, respectively. The neutral GAG’s favors the digestive transit, lubricates food particles, allows them to be pushed to the next segment. The acid GAG’s are very important to trap particles and form a protective barrier against mechanical friction and the action of the digestive acid [3]. The beginning of the secretions which had positive reaction to AB and PAS staining was noted in the pheasant thoracic esophagus on the 21st day of incubation period and a day later in the cervical region. These observations are reported 3 days earlier and 2 days later in the thoracic and cervical regions of partridge esophagus, respectively [17].
The primordial glandular tissue of Gallus gallus domesticus was weakly positive to AB and PAS staining in the last quarter of embryonic period. However, strong positive reaction to these staining in the next days showed full production of neutral and acid GAG’s in these species [3]. In the guinea fowl, near the end of the incubation period, the epithelium and primordial glands demonstrated weak reactions to PAS. However, the glands show mucosal characteristics at the end of incubation period [16].
In conclusion, the this study declared that the most esophageal developmental events of the pheasant occur in the first 2-thirds of the incubation period. Most of these events were observed in the thoracic esophagus earlier than cervical region. Comparison of our results with similar studieson the avian esophagus development, indicates that the sequence of the developmental events in pheasant are similar to the other birds; however, the three are variations in onset time of events. The differentiation of the epithelial cells into stratified squamous occur in the initial days of second half of pheasant incubation period., but However, it is seen at the last third stage in partridge and guinea fowl.
Cilia appear in the initial day of pheasant and partridge embryonic period. However, previous studies reported its occurrence almost in the middle or near the end of incubation period. The esophageal glands of pheasant and partridge form at the last third of incubation time, but they appear almost in the second half of chicken and middle third of turkey embryonic period. According to this study and the other investigations, the formation of lumen of glands is obsereved near the end of incubation. However, it occurs in the middle of the embryonic period among Gallus gallus domesticus.
Some of these variations can be related to the differences in the length of the incubation period in different species. In this case, a correlation is expected between the variations of developmental events and the length of incubation periods. The present study indicates that this prospect works for pheasant and partridge as 2 members of Phasianidae. However, this rule is not reported regarding other studied species.
Ethical Considerations
Compliance with ethical guidelines
All steps of this study were approved by the Animal Ethics Committee, based on the Research Council of the Veterinary Faculty of Tehran University.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Authors contributions
All authors have read and approved the manuscript.
Conflict of interest
The authors declared no conflict of interest.
Acknowledgments
This research was financially supported by the Research Council of Faculty of Veterinary Medicine, University of Tehran.
References
- Narita T, Saitoh K, Kameda T, Kuroiwa A, Mizutani M, Koike C, et al. BMPs are necessary for stomach gland formation in the chicken embryo: A study using virally induced BMP-2 and Noggin expression. Development. 2000; 127(5):981-8. [PMID]
- Allenspach AL. The reopening process of the esophagus in the normal chick and the crookedneck dwarf mutant. Development. 1966; 15(1):67-76.
- Ventura A, do Nascimento AA, dos Santos MAJ, Vieira-Lopes DA, Sales A, Pinheiro NL. Histological description of morphogenesis of the gastroesophageal mucosa of gallus gallus domesticus (Linnaeus, 1758). International Journal of Morphology. 2013; 31(4):1331-9. [DOI:10.4067/S0717-95022013000400030]
- Wakely J, England MA. Scanning electron microscopical and histochemical study of the structure and function of basement membranes in the early chick embryo. Proceedings of the Royal Society of London. 1979; 206(1164):329-52. [DOI:10.1098/rspb.1979.0109] [PMID]
- Bellairs R, Osmond M. Atlas of chick development. Amesterdam: Elsevier; 2005. [PMID]
- Bacha Jr WJ, Bacha LM. Color atlas of veterinary histology. Hoboken, New Jersey: John Wiley & Sons; 2012.
- Banks WJ. Applied veterinary histology. Maryland Heights, Missouri: Mosby; 1993.
- Ivey W, Edgar S. The histogenesis of the esophagus and crop of the chicken, turkey, guinea fowl and pigeon, with special reference to ciliated epithelium. The Anatomical Record. 1952; 114(2):189-211. [DOI:10.1002/ar.1091140207] [PMID]
- Parchami A, Dehkordi R. Histological characteristics of the esophageal wall of the common Quail. World Applied Sciences Journal. 2011; 14(3):414-9.
- Sağsöz H, Lıman N. Structure of the oesophagus and morphometric, histochemical–immunohistochemical profiles of the oesophageal gland during the post‐hatching period of Japanese Quails (coturnix coturnix Japonica). Anatomia, Histologia, Embryologia. 2009; 38(5):330-40. [DOI:10.1111/j.1439-0264.2009.00947.x] [PMID]
- Rossi JR, Baraldi-Artoni SM, Oliveira D, Da Cruz C, Sagula A, Pacheco MR, et al. Morphology of oesophagus and crop of the partrigde Rhynchotus rufescens (Tiramidae). Acta Scientiarum: Biological Sciences. 2006; 28(2):165-8.
- Mobini B. The effect of age, sex and region on histological structures of the esophagus in broiler chickens. Veterinarija ir Zootechnika. 2014; 66(88):46-9.
- Madhu N, Balasundaram K, Paramasivan S, Jayachitra S, Vijayakumar K, Tamilselvan S. Gross morphology and histology of oesophagus in adult emu birds (dromaius novaehollandiae). Asian Journal of Science and Technology. 2015; 6(1):969-71.
- Rajabi E, Nabipour A. Histological study on the oesophagus and crop in various species of wild bird. Avian Biology Research. 2009; 2(3):161-4. [DOI:10.3184/175815509X12474789336122]
- Shehan N. Anatomical and histological study of esophagus in Geese (Anser anser demesticus). Basrah Journal Veterinary Research. 2012; 11(1):13-22.
- Gosomji IJ, Salami SO, Nzalak JO, Kawu MU, Tizhe EV, Gurumyen YG, et al. Histogenesis of the oesophagus of guinea fowl (numida meleagris) at prehatch and posthatch. Scientifica. 2016; Article ID:9827956.
- Shojaei B, Hashemnia S, Rad RE. Histochemical study of the oesophagus in the chukar partridge (Alectoris chukar) embryo. Folia Morphologica. 2016; 75(4):474-80. [DOI:10.5603/FM.a2016.0011] [PMID]
- McGeady TA, Quinn PJ, FitzPatrick E, Ryan M, Kilroy D, Lonergan P. Veterinary embryology. Hoboken, New Jersey: John Wiley & Sons; 2017.
- Luna LG. Manual of histologic staining methods of the Armed Forces Institute of Pathology. Rome: Food and Agriculture Organization; 1968.
- Whitehead RH. A note on the development of the esophageal epithelium. American Journal of Anatomy. 1905; 4:vi-vii.
- Johnson FP. The development of the mucous membrane of the oesophagus, stomach and small intestine in the human embryo. Developmental Dynamics. 1910; 10(1):521-75.
- Raymond C, Anne V, Millane G. Development of esophageal epithelium in the fetal and neonatal mouse. The Anatomical Record. 1991; 230(2):225-34. [DOI:10.1002/ar.1092300210] [PMID]
- Rodrigues MN, Carvalho RC, Franciolli AL, Rodrigues RF, Rigoglio NN, Jacob JC, et al. Prenatal development of the digestive system in the horse. The Anatomical Record. 2014; 297(7):1218-27. [DOI:10.1002/ar.22929] [PMID]
- Bello A, Onyeanusi B, Sonfada M, Adeyanju J, Umar A, Umaru M, et al. Histomorphological studies of the prenatal development of oesophagus of one humped camel (camelus dromedarius). Scientific Journal of Agriculture. 2012; 1(4):100-4.
- Panchamukhi BG. Prenatal development of the buffalo babalus bubalis stomach with particular reference to organogenesis and histogenesis [PhD dissertation]. Gujarat: Sardar Patel University; 1973.
- Drummond IA. Cilia functions in development. Current Opinion in Cell Biology. 2012; 24(1):24-30. [DOI:10.1016/j.ceb.2011.12.007] [PMID] [PMCID]
- Engvall E. Structure and function of basement membranes. International Journal of Developmental Biology. 2003; 39(5):781-7. [PMID]
Type of Study: Original |
Subject:
Developmental Anatomy
Received: 2018/03/10 | Accepted: 2018/11/2 | Published: 2019/01/1
Received: 2018/03/10 | Accepted: 2018/11/2 | Published: 2019/01/1
References
1. Narita T, Saitoh K, Kameda T, Kuroiwa A, Mizutani M, Koike C, et al. BMPs are necessary for stomach gland formation in the chicken embryo: A study using virally induced BMP-2 and Noggin expression. Development. 2000; 127(5):981-8. [PMID] [PMID]
2. Allenspach AL. The reopening process of the esophagus in the normal chick and the crookedneck dwarf mutant. Development. 1966; 15(1):67-76.
3. Ventura A, do Nascimento AA, dos Santos MAJ, Vieira-Lopes DA, Sales A, Pinheiro NL. Histological description of morphogenesis of the gastroesophageal mucosa of gallus gallus domesticus (Linnaeus, 1758). International Journal of Morphology. 2013; 31(4):1331-9. [DOI:10.4067/S0717-95022013000400030] [DOI:10.4067/S0717-95022013000400030]
4. Wakely J, England MA. Scanning electron microscopical and histochemical study of the structure and function of basement membranes in the early chick embryo. Proceedings of the Royal Society of London. 1979; 206(1164):329-52. [DOI:10.1098/rspb.1979.0109] [PMID] [DOI:10.1098/rspb.1979.0109]
5. Bellairs R, Osmond M. Atlas of chick development. Amesterdam: Elsevier; 2005. [PMID] [PMID]
6. Bacha Jr WJ, Bacha LM. Color atlas of veterinary histology. Hoboken, New Jersey: John Wiley & Sons; 2012.
7. Banks WJ. Applied veterinary histology. Maryland Heights, Missouri: Mosby; 1993.
8. Ivey W, Edgar S. The histogenesis of the esophagus and crop of the chicken, turkey, guinea fowl and pigeon, with special reference to ciliated epithelium. The Anatomical Record. 1952; 114(2):189-211. [DOI:10.1002/ar.1091140207] [PMID] [DOI:10.1002/ar.1091140207]
9. Parchami A, Dehkordi R. Histological characteristics of the esophageal wall of the common Quail. World Applied Sciences Journal. 2011; 14(3):414-9.
10. Sağsöz H, Lıman N. Structure of the oesophagus and morphometric, histochemical–immunohistochemical profiles of the oesophageal gland during the post‐hatching period of Japanese Quails (coturnix coturnix Japonica). Anatomia, Histologia, Embryologia. 2009; 38(5):330-40. [DOI:10.1111/j.1439-0264.2009.00947.x] [PMID] [DOI:10.1111/j.1439-0264.2009.00947.x]
11. Rossi JR, Baraldi-Artoni SM, Oliveira D, Da Cruz C, Sagula A, Pacheco MR, et al. Morphology of oesophagus and crop of the partrigde Rhynchotus rufescens (Tiramidae). Acta Scientiarum: Biological Sciences. 2006; 28(2):165-8.
12. Mobini B. The effect of age, sex and region on histological structures of the esophagus in broiler chickens. Veterinarija ir Zootechnika. 2014; 66(88):46-9.
13. Madhu N, Balasundaram K, Paramasivan S, Jayachitra S, Vijayakumar K, Tamilselvan S. Gross morphology and histology of oesophagus in adult emu birds (dromaius novaehollandiae). Asian Journal of Science and Technology. 2015; 6(1):969-71.
14. Rajabi E, Nabipour A. Histological study on the oesophagus and crop in various species of wild bird. Avian Biology Research. 2009; 2(3):161-4. [DOI:10.3184/175815509X12474789336122] [DOI:10.3184/175815509X12474789336122]
15. Shehan N. Anatomical and histological study of esophagus in Geese (Anser anser demesticus). Basrah Journal Veterinary Research. 2012; 11(1):13-22.
16. Gosomji IJ, Salami SO, Nzalak JO, Kawu MU, Tizhe EV, Gurumyen YG, et al. Histogenesis of the oesophagus of guinea fowl (numida meleagris) at prehatch and posthatch. Scientifica. 2016; Article ID:9827956.
17. Shojaei B, Hashemnia S, Rad RE. Histochemical study of the oesophagus in the chukar partridge (Alectoris chukar) embryo. Folia Morphologica. 2016; 75(4):474-80. [DOI:10.5603/FM.a2016.0011] [PMID] [DOI:10.5603/FM.a2016.0011]
18. McGeady TA, Quinn PJ, FitzPatrick E, Ryan M, Kilroy D, Lonergan P. Veterinary embryology. Hoboken, New Jersey: John Wiley & Sons; 2017.
19. Luna LG. Manual of histologic staining methods of the Armed Forces Institute of Pathology. Rome: Food and Agriculture Organization; 1968.
20. Whitehead RH. A note on the development of the esophageal epithelium. American Journal of Anatomy. 1905; 4:vi-vii.
21. Johnson FP. The development of the mucous membrane of the oesophagus, stomach and small intestine in the human embryo. Developmental Dynamics. 1910; 10(1):521-75.
22. Raymond C, Anne V, Millane G. Development of esophageal epithelium in the fetal and neonatal mouse. The Anatomical Record. 1991; 230(2):225-34. [DOI:10.1002/ar.1092300210] [PMID] [DOI:10.1002/ar.1092300210]
23. Rodrigues MN, Carvalho RC, Franciolli AL, Rodrigues RF, Rigoglio NN, Jacob JC, et al. Prenatal development of the digestive system in the horse. The Anatomical Record. 2014; 297(7):1218-27. [DOI:10.1002/ar.22929] [PMID] [DOI:10.1002/ar.22929]
24. Bello A, Onyeanusi B, Sonfada M, Adeyanju J, Umar A, Umaru M, et al. Histomorphological studies of the prenatal development of oesophagus of one humped camel (camelus dromedarius). Scientific Journal of Agriculture. 2012; 1(4):100-4.
25. Panchamukhi BG. Prenatal development of the buffalo babalus bubalis stomach with particular reference to organogenesis and histogenesis [PhD dissertation]. Gujarat: Sardar Patel University; 1973.
26. Drummond IA. Cilia functions in development. Current Opinion in Cell Biology. 2012; 24(1):24-30. [DOI:10.1016/j.ceb.2011.12.007] [PMID] [PMCID] [DOI:10.1016/j.ceb.2011.12.007]
27. Engvall E. Structure and function of basement membranes. International Journal of Developmental Biology. 2003; 39(5):781-7. [PMID]
Send email to the article author
| Rights and permissions | |
 |
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License. |

Copyright © The Author(s);
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC-By-NC), which permits use, distribution, and reproduction in any medium, provided the original work is properly cited and is not used for commercial purposes.
Contact Information
